N.B: Petite vidéo que j'avais visionné au nouvel an dernier et qu'avait posté la connaissance d'un forum.
samedi 31 décembre 2011
lundi 26 décembre 2011
L'importance méconnue de l'évolution neutre
Ce présent message reprend en grande partie un message que j’avais posté sur un forum de discussion sous le pseudo de uno. Je me suis en effet dit qu’il serait intéressant de revenir sur cet exemple précis qu’est celui de l’évolution chez le Rat-Taupe du gène codant la protéine «αA Cristalline» et de ses application en matière de compréhension de cette discipline scientifique fascinante qu’est la biologie de l’évolution.
Lorsque les gens entendent parler de la Théorie de l’évolution, ils pensent généralement immédiatement à Charles Robert Darwin, à la sélection naturelle et à la survie des plus aptes. Pire beaucoup de gens confondent évolution et adaptation!
Or bien évidemment l’écrasante majorité des biologistes (à l’exception de quelques «ultra-darwiniens») ne tombent pas dans ce travers et savent que si l’adaptation est une composante importante de l’évolution biologique, elle n’en est de loin pas la seule et ne peut donc être ainsi confondu avec l’évolution.
Par exemple le biochimiste Laurence A. Moran a rappelé a mentionné à plusieurs reprises sur son blog l’importance de la dérive génétique (bien que récemment on m'a fait remarqué qu'il avait tendance à appeler «dérive génétique» tout et n'importe quoi) et plus généralement de l’évolution neutre dans l’évolution des êtres vivants.
Pour rappelle la dérive génétique est grossomodo le changement de fréquences allélique au sein d’une population via «le tirage au sort des allèles» lors de la reproduction des individus et donc la dérive génétique n’implique pas de sélection favorisant tel ou tel allèle par apport à un autre. L’évolution neutre conceptualisée par le généticien japonais Motoo Kimura, reprend notamment le concept de «dérive génétique» mais en se focalisant sur les mutations elles-mêmes. C'est-à-dire en soulignant que de nombreuses mutations sont sans conséquences en soulignant que celles-ci sont neutre du point de vue sélectif pourles organisme. C’est ainsi que le paradigme de l’évolution neutre aussi parfois appelé «Théorie neutraliste de l’évolution» en arrive à la conclusion que de nombreux changements affectant l’évolution de notre génome, sont donc non-adaptatifs.

Et c’est là que la théorie neutraliste de l’évolution devient intéressante, car donc celle-ci souligne également que les séquences d’ADN de notre génome remplissant une fonctionnalité importantes accumulent donc moins de mutations que celles qui demeurent inutiles. Pourquoi? Simplement parce que les mutations touchant des parties fonctionnelles de notre génome, endommageront, dans une certaine fréquence, celles-ci et seront donc éliminées par des balayages sélectifs. Certes il est important de souligner que la théorie de Kimura inclue également des mutations touchant des régions fonctionnelles de notre génome mais qui n'ont elles-mêmes pas d'impact en terme de valeur sélective et donc entre également dans la théorie de l'évolution neutre de Motoo Kimura.* Cependant les régions non-fonctionnels (ou peu fonctionnel) de notre génome accumuleront donc et donc davantage de mutations. Cela veut dire que plus une partie de notre génome est dépourvue de fonctionnalité plus celle-ci tendra à accumuler davantage de mutations.
À ce titre je reviens sur un exemple très illustratif, celui d’une séquence génétique codant une protéine nommé «αA Cristalline» qui a été étudié chez différentes espèces de rongeurs [1]. En effet une équipe de chercheur a étudié la séquence d’ADN codant cette protéine chez différentes rongeurs à savoir une espèce de Castor, le Goundi de l'Atlas, l’Écureuil roux, une espèce de Rat, une autre souris, un Hamster, et comme nous le verrons l'espèce la plus intéressante à savoir le Rat taupe (Spalax ehrenbergi). Parmi ces espèces il y en a une qui est aveugle et c’est bien évidemment le Rat Taupe nommé Spalax ehrenbergi.

Or chose intéressante la protéine «αA Cristalline» est, comme certains l'on peut-être déjà deviné via son nom, impliqué dans la vue. C’est à partir de là que l’on peut faire une intéressante hypothèse. Sachant que cette protéine accomplit une fonction cruciale dans la vision des animaux il est possible ou tout du moins probable que le gène codant la protéine «αA Cristalline» soit «conservée» c’est-à-dire soumises à de nombreux «balayages sélectifs» éliminant la plupart des mutations et donc que cette séquence évolue relativement lentement au fil du temps. Cependant cela ne serait vrai que si la protéine en question doit être «conservée» en vue de son rôle crucial dans la vision. Car chez le Rat Taupe, animal aveugle, cette protéine n’a plus ce rôle prépondérant. Et donc contrairement aux autres rongeurs le gène codant la protéine «αA Cristalline» a probablement accumulé d’avantage de mutations et donc évolué plus rapidement que chez les autres rongeurs ayant encore une vision normale.
Ainsi c’est dans cette optique qu’une équipe de chercheur a étudié la séquence du gène codant la protéine «αA Cristalline» chez différentes espèces de rongeurs à vision efficiente avec celle du Rat Taupe. Et les résultats de l’étude sont édifiants comme l’a retranscrit Stephen Jay Gould.
Ainsi la même séquence peut évoluer un rythme différents dans différentes lignée. La séquence codant la protéine «αA Cristalline» ayant évolué plus rapidement chez Spalax ehrenbergi que chez d’autres lignées de rongeurs et plus vite que chez bien d’autres lignées encore.
 Ce schéma tiré de l’étude portant sur l’évolution de la protéine «αA Cristalline» chez Spalax ehrenbergi, montre les changements survenus dans la dite protéine «αA Cristalline» au sein de différentes lignées de mammifères! On remarque que si la lignée des Castors (Beaver en anglais) a accumulé elle aussi quelque changements, la lignée des Rats-Taupes (Mole Rat en anglais) a vu son «αA Cristalline» accumulé un nombre inégalé de changements par-apport aux autres lignées. Cela confirmant l’hypothèse selon laquelle un relâchement de la sélection naturelle pour «l’αA Cristalline» dans la lignée du Rat-Taupe a permis à la séquence d’ADN codant cette protéine d’évolué beaucoup plus rapidement c’est-à-dire d’accumuler un très grande nombre de mutations.
Ce schéma tiré de l’étude portant sur l’évolution de la protéine «αA Cristalline» chez Spalax ehrenbergi, montre les changements survenus dans la dite protéine «αA Cristalline» au sein de différentes lignées de mammifères! On remarque que si la lignée des Castors (Beaver en anglais) a accumulé elle aussi quelque changements, la lignée des Rats-Taupes (Mole Rat en anglais) a vu son «αA Cristalline» accumulé un nombre inégalé de changements par-apport aux autres lignées. Cela confirmant l’hypothèse selon laquelle un relâchement de la sélection naturelle pour «l’αA Cristalline» dans la lignée du Rat-Taupe a permis à la séquence d’ADN codant cette protéine d’évolué beaucoup plus rapidement c’est-à-dire d’accumuler un très grande nombre de mutations.
Certains pourraient rétorquer que certes, l’évolution neutre a son importance, mais cette importance se limite au seul niveau moléculaire, c’est-à-dire ne concerne que des séquences d’ADN dont la modification n’a pas de conséquences phénotypiques et donc adaptatives, notables.
Mais cela est faux! Cela est faux d’une part parce que des caractéristiques phénotypiques peuvent également très facilement se répandre par la simple dérive génétique. Mais aussi et peut-être même surtout, parce que l’évolution neutre peut avoir un grand impact sur l’évolution adaptative elle-même!
J’avais en effet rappelé dans ce précédent message répondant à quelques inepties créationnistes, en quoi l’évolution neutre pouvait avoir son importance dans l’apparition de nouvelles adaptations. J’avais notamment illustré cela via le schéma suivant.
 Soit un séquence d’ADN fonctionnel perdant sa fonctionnalité suite à une première mutation (1). La perte de fonctionnalité pouvant être sans conséquence négative pour la survie de l’organisme concernée, celui-ci pourra donc continué à se reproduire. Mais donc cette séquence d’ADN devenu non-fonctionnelle fera qu’au fil des générations celle-ci pourra accumuler librement un grand nombre de mutations comme le stipule la théorie neutraliste de l’évolution (2) (3) (4). Or il n’est pas exclu qu’à terme certaines des mutations accumulés ouvrent la voie à l’apparition d’une nouvelle fonctionnalité pour la séquence d’ADN en question (5).
Soit un séquence d’ADN fonctionnel perdant sa fonctionnalité suite à une première mutation (1). La perte de fonctionnalité pouvant être sans conséquence négative pour la survie de l’organisme concernée, celui-ci pourra donc continué à se reproduire. Mais donc cette séquence d’ADN devenu non-fonctionnelle fera qu’au fil des générations celle-ci pourra accumuler librement un grand nombre de mutations comme le stipule la théorie neutraliste de l’évolution (2) (3) (4). Or il n’est pas exclu qu’à terme certaines des mutations accumulés ouvrent la voie à l’apparition d’une nouvelle fonctionnalité pour la séquence d’ADN en question (5).
Le fait que certaines mutations neutres sur le plan sélectif puissent constituer des «étapes» préalables à l’apparition de nouvelles adaptations a en effet fait l’objet des publications scientifiques très intéressantes ces dernières années. Nous pouvons par exemple cité les travaux du généticien Michael Lynch [2], ou encore ceux de Joseph W. Thornton sur l’évolution d’un récepteur hormonal notamment via là aussi l’évolution neutre [3]. Enfin les expériences d’évolution en laboratoire de souches de la bactérie Escherichia coli ont également montré comment des mutations neutres apparus au préalables sans raison adaptative peuvent néanmoins ouvrir la voie à de nouvelles adaptations à venir. Ces travaux soulignant en quoi l’évolution neutre peut influer sur l’apparition de possibles adaptations [4] [5].
En conclusion il apparait donc clairement qu’il est important de prendre en compte l’importance considérable de l’évolution neutre pour comprendre comme il se doit l’évolution en général. Car si la sélection naturelle conceptualisé en son temps par Charles Robert Darwin est sans aucun doute une composante importante de l'évolution, elle n’en est pas moins insuffisante pour comprendre la dite évolution. L’évolution neutre nous amène à nous rendre compte que l’évolution même si soumise au déterminisme de la sélection naturelle n’en demeure pas moins essentiellement contingente. Voici un point sur lequel Stephen Jay Gould avait très probablement vu on ne peut pas plus juste.
* Oui car comme on me l'a rappelé récemment l'évolution neutre inclue également des mutations touchant des régions fonctionnels de notre génome, y compris des régions codantes. Il s'agit simplement de mutation n'impactant pas négativement ou positivement la séquence concernée.
Références:
[1] Wiljan Hendriks et al (1987), The lens protein aA-crystallin of the blind mole rat, Spalax ehrenbergi: Evolutionary change and functional constraints, Proceedings of the National Academy of Sciences
[2] Michael Lynch (2010), Scaling expectations for the time to establishment of complex adaptations, Proceedings of the National Academy of Sciences
[3] Jamie T. Bridgham, Eric A. Ortlund and Joseph W. Thornton (2009), An epistatic ratchet constrains the direction of glucocorticoid receptor evolution, Nature
[4] Jeffrey E. Barrick et al (2010), Escherichia coli rpoB Mutants Have Increased Evolvability in Proportion to Their Fitness Defects, Molecular Biology and Evolution
[5] Robert J. Woods et al (2011), Second-Order Selection for Evolvability in a Large Escherichia coli Population, Science
Lorsque les gens entendent parler de la Théorie de l’évolution, ils pensent généralement immédiatement à Charles Robert Darwin, à la sélection naturelle et à la survie des plus aptes. Pire beaucoup de gens confondent évolution et adaptation!
Or bien évidemment l’écrasante majorité des biologistes (à l’exception de quelques «ultra-darwiniens») ne tombent pas dans ce travers et savent que si l’adaptation est une composante importante de l’évolution biologique, elle n’en est de loin pas la seule et ne peut donc être ainsi confondu avec l’évolution.
Par exemple le biochimiste Laurence A. Moran a rappelé a mentionné à plusieurs reprises sur son blog l’importance de la dérive génétique (bien que récemment on m'a fait remarqué qu'il avait tendance à appeler «dérive génétique» tout et n'importe quoi) et plus généralement de l’évolution neutre dans l’évolution des êtres vivants.
Pour rappelle la dérive génétique est grossomodo le changement de fréquences allélique au sein d’une population via «le tirage au sort des allèles» lors de la reproduction des individus et donc la dérive génétique n’implique pas de sélection favorisant tel ou tel allèle par apport à un autre. L’évolution neutre conceptualisée par le généticien japonais Motoo Kimura, reprend notamment le concept de «dérive génétique» mais en se focalisant sur les mutations elles-mêmes. C'est-à-dire en soulignant que de nombreuses mutations sont sans conséquences en soulignant que celles-ci sont neutre du point de vue sélectif pourles organisme. C’est ainsi que le paradigme de l’évolution neutre aussi parfois appelé «Théorie neutraliste de l’évolution» en arrive à la conclusion que de nombreux changements affectant l’évolution de notre génome, sont donc non-adaptatifs.
Le généticien Motoo Kimura père de la théorie neutraliste de l'évolution

Et c’est là que la théorie neutraliste de l’évolution devient intéressante, car donc celle-ci souligne également que les séquences d’ADN de notre génome remplissant une fonctionnalité importantes accumulent donc moins de mutations que celles qui demeurent inutiles. Pourquoi? Simplement parce que les mutations touchant des parties fonctionnelles de notre génome, endommageront, dans une certaine fréquence, celles-ci et seront donc éliminées par des balayages sélectifs. Certes il est important de souligner que la théorie de Kimura inclue également des mutations touchant des régions fonctionnelles de notre génome mais qui n'ont elles-mêmes pas d'impact en terme de valeur sélective et donc entre également dans la théorie de l'évolution neutre de Motoo Kimura.* Cependant les régions non-fonctionnels (ou peu fonctionnel) de notre génome accumuleront donc et donc davantage de mutations. Cela veut dire que plus une partie de notre génome est dépourvue de fonctionnalité plus celle-ci tendra à accumuler davantage de mutations.
À ce titre je reviens sur un exemple très illustratif, celui d’une séquence génétique codant une protéine nommé «αA Cristalline» qui a été étudié chez différentes espèces de rongeurs [1]. En effet une équipe de chercheur a étudié la séquence d’ADN codant cette protéine chez différentes rongeurs à savoir une espèce de Castor, le Goundi de l'Atlas, l’Écureuil roux, une espèce de Rat, une autre souris, un Hamster, et comme nous le verrons l'espèce la plus intéressante à savoir le Rat taupe (Spalax ehrenbergi). Parmi ces espèces il y en a une qui est aveugle et c’est bien évidemment le Rat Taupe nommé Spalax ehrenbergi.
Photo de Rat Taupe

Or chose intéressante la protéine «αA Cristalline» est, comme certains l'on peut-être déjà deviné via son nom, impliqué dans la vue. C’est à partir de là que l’on peut faire une intéressante hypothèse. Sachant que cette protéine accomplit une fonction cruciale dans la vision des animaux il est possible ou tout du moins probable que le gène codant la protéine «αA Cristalline» soit «conservée» c’est-à-dire soumises à de nombreux «balayages sélectifs» éliminant la plupart des mutations et donc que cette séquence évolue relativement lentement au fil du temps. Cependant cela ne serait vrai que si la protéine en question doit être «conservée» en vue de son rôle crucial dans la vision. Car chez le Rat Taupe, animal aveugle, cette protéine n’a plus ce rôle prépondérant. Et donc contrairement aux autres rongeurs le gène codant la protéine «αA Cristalline» a probablement accumulé d’avantage de mutations et donc évolué plus rapidement que chez les autres rongeurs ayant encore une vision normale.
Ainsi c’est dans cette optique qu’une équipe de chercheur a étudié la séquence du gène codant la protéine «αA Cristalline» chez différentes espèces de rongeurs à vision efficiente avec celle du Rat Taupe. Et les résultats de l’étude sont édifiants comme l’a retranscrit Stephen Jay Gould.
«La protéine codée par le gène en question chez Spalax, par exemple, a subit le remplacement de neuf acides aminés (sur 173 changements possibles) par rapport à l’état ancestral de la protéine dans ce groupe (il s’agit de la famille des muridés comprenant les rats, les souris, le hamster et la gerbille) présentent des séquences identiques, n’ayant subi absolument aucune changement par apport à l’état ancestral. On a établi que, dans l’ensemble des vertébrés, le rythme moyen de changement dans l’αA cristalline est d’environ 3 remplacements d’acides aminés par 100 positions par 100 millions d’années. Spalax est en train de changer à un rythme plus de quatre fois supérieur, soit environ 13% pour cent millions d’années. (Neuf changements sur 173 positions représentent 5,2%; mais la lignée de Spalax n'a que 40 millions d'années et 5,2% en quarante millions d’années correspond à 13% en 100 millions d’années.) En outre, Spalax a changé quatre acides aminés au niveau de positions qui sont absolument invariables chez tous les autres vertébrés étudiés - lesquels représentent soixante-douze espèces, allant du requin appelé «roussette» jusqu’à l’espèce humaine.» Stephen Jay Gould, Comme les huit doigts de la main, Éditions du Seuil 1996
Ainsi la même séquence peut évoluer un rythme différents dans différentes lignée. La séquence codant la protéine «αA Cristalline» ayant évolué plus rapidement chez Spalax ehrenbergi que chez d’autres lignées de rongeurs et plus vite que chez bien d’autres lignées encore.

Certains pourraient rétorquer que certes, l’évolution neutre a son importance, mais cette importance se limite au seul niveau moléculaire, c’est-à-dire ne concerne que des séquences d’ADN dont la modification n’a pas de conséquences phénotypiques et donc adaptatives, notables.
Mais cela est faux! Cela est faux d’une part parce que des caractéristiques phénotypiques peuvent également très facilement se répandre par la simple dérive génétique. Mais aussi et peut-être même surtout, parce que l’évolution neutre peut avoir un grand impact sur l’évolution adaptative elle-même!
J’avais en effet rappelé dans ce précédent message répondant à quelques inepties créationnistes, en quoi l’évolution neutre pouvait avoir son importance dans l’apparition de nouvelles adaptations. J’avais notamment illustré cela via le schéma suivant.

Le fait que certaines mutations neutres sur le plan sélectif puissent constituer des «étapes» préalables à l’apparition de nouvelles adaptations a en effet fait l’objet des publications scientifiques très intéressantes ces dernières années. Nous pouvons par exemple cité les travaux du généticien Michael Lynch [2], ou encore ceux de Joseph W. Thornton sur l’évolution d’un récepteur hormonal notamment via là aussi l’évolution neutre [3]. Enfin les expériences d’évolution en laboratoire de souches de la bactérie Escherichia coli ont également montré comment des mutations neutres apparus au préalables sans raison adaptative peuvent néanmoins ouvrir la voie à de nouvelles adaptations à venir. Ces travaux soulignant en quoi l’évolution neutre peut influer sur l’apparition de possibles adaptations [4] [5].
En conclusion il apparait donc clairement qu’il est important de prendre en compte l’importance considérable de l’évolution neutre pour comprendre comme il se doit l’évolution en général. Car si la sélection naturelle conceptualisé en son temps par Charles Robert Darwin est sans aucun doute une composante importante de l'évolution, elle n’en est pas moins insuffisante pour comprendre la dite évolution. L’évolution neutre nous amène à nous rendre compte que l’évolution même si soumise au déterminisme de la sélection naturelle n’en demeure pas moins essentiellement contingente. Voici un point sur lequel Stephen Jay Gould avait très probablement vu on ne peut pas plus juste.
* Oui car comme on me l'a rappelé récemment l'évolution neutre inclue également des mutations touchant des régions fonctionnels de notre génome, y compris des régions codantes. Il s'agit simplement de mutation n'impactant pas négativement ou positivement la séquence concernée.
Références:
[1] Wiljan Hendriks et al (1987), The lens protein aA-crystallin of the blind mole rat, Spalax ehrenbergi: Evolutionary change and functional constraints, Proceedings of the National Academy of Sciences
[2] Michael Lynch (2010), Scaling expectations for the time to establishment of complex adaptations, Proceedings of the National Academy of Sciences
[3] Jamie T. Bridgham, Eric A. Ortlund and Joseph W. Thornton (2009), An epistatic ratchet constrains the direction of glucocorticoid receptor evolution, Nature
[4] Jeffrey E. Barrick et al (2010), Escherichia coli rpoB Mutants Have Increased Evolvability in Proportion to Their Fitness Defects, Molecular Biology and Evolution
[5] Robert J. Woods et al (2011), Second-Order Selection for Evolvability in a Large Escherichia coli Population, Science
Le retour des BANDits
Mon petit dossier en cinq parties (parties 1, 2, 3, 4, 5) sur les BANDits («Birds Are Not Dinosaurs» minorité de chercheurs niant l’origine dinosaurienne des oiseaux) peut toujours s’avérer utile comme référence vis-à-vis des quelques zigotos créationnistes ou non, qui s’acharnent mordicus à nier que oiseaux sont des dinosaures en dépit des multiples preuves et démonstrations accumulé depuis plusieurs décennies déjà. À ce titre notons qu’Alan Feduccia, un célèbre BANDit vient de pondre un livre qui sera sauf erreur disponible dès janvier 2012. La description de ce livre donne le ton à lui seul.
«En examinant et interprétant les récentes et spectaculaires découvertes fossiles en Chine, les paléontologues sont arrivés à un point de vue prévalant: « Il y a maintenant des preuves irréfutables que les oiseaux représentent la dernière lignée de dinosaures, encore vivante. Mais c'est à cette conclusion est-elle incontestable? Dans ce livre, le biologiste évolutionniste Alan Feduccia fournit l'analyse la plus compréhensive à ce jour, des oiseaux et autres preuves et associés trouvés en Chine, il expose ensuite les spéculations massives sans fondement qui ont accompagné ces découvertes et qui ont été publiées dans les pages de revues scientifiques prestigieuses.»
«Alan Feduccia soutient que les défenseurs de l'orthodoxie actuelle sur les origines des oiseaux ont ignoré les données contraires, mal-interprété les fossiles, et utilisé un raisonnement erroné. Il examine pourquoi et comment le débat est devenu tellement polémique et s’adonne à un plaidoyer pour recentrer la discussion en "rompant avec des camisoles de force méthodologique et voir le monde des origines d’un œil nouveau." S'appuyant sur une vie d'étude, il propose sa propre compréhension actuelle de l’origine et du vol des oiseaux.»
Alan Feduccia compte donc clairement continuer à nier l’origine dinosaurienne des oiseaux en accusant la majorité des chercheurs travaillant sur la question de l’origine évolutive des oiseaux d’être au mieux des simplets incompétents au pire de méchants dogmaticiens s’opposant à la «vérité qui dérange» dont Alan Feduccia et les autres BANDits en seraient les détenteurs éclairés.
Tout cela ne tient bien évidemment pas, car nous l’avons vu en long et en large Alan Feduccia et les autres BANDits sont justement ceux dont le raisonnement est erroné et qui ignorent les preuves s’opposant à leurs idées. Alan Feduccia s’est même vu totalement contredire et pire encore s’est lui-même auto-contredit sans jamais admettre comme il se devait ses torts. Le pauvre Alan Feduccia qui affirmait jusqu’au début des années 2000 que les Dromaeosauridés tels que le célèbre Vélociraptor ne peuvent être apparenté aux oiseaux à cause de leurs dents et de leurs crânes trop dinosauriens à son goût avant d’affirmer dans les années 2000 suite à la découverte Dromaeosauridés à plumes que ces mêmes Dromaeosauridés sont des oiseaux et non pas des dinosaures…..On peut résumer «l’argumentation d’Alan Feduccia» via l’image suivante.

Désolé mon cher Alan Feduccia mais tous les livres que vous publierez ne pourront pas effacer vos auto-contradictions et le caractère bancale et transpirant la mauvaise foi de vos affirmations car vous ne faites qu’accuser les autres chercheurs de choses dont seuls vous et les autres BANDits faites preuves.
dimanche 25 décembre 2011
lundi 5 décembre 2011
Erreurs et correction de « L'origine évolutive d'étranges insectes »

Le mois de Mai dernier j'avais posté un message consacré à l'évolution d'étranges insectes à savoir les Membracides.
Ces message intitulée L'origine évolutive d'étranges insectes contenait néanmoins deux erreurs qu'une connaissance du site rationalisme.org (site figurant dorénavant à la liste des sites recommandés) m'a signalé et mieux encore corrigé.
La correction en question figure au bas de mon message consacré aux Membracides.
Un grand merci à Yenagna Landré pour cette correction!
mercredi 2 novembre 2011
L'incertaine position phylogénétique d'Archaeoptéryx

Dans un de mes précédemment message j’étais revenu sur la découverte d’un nouveau dinosaure à plumes nommé Xiaotingia zhengi et des implications de celui-ci en matière de phylogénie, notamment en ce qui concerne le très célèbre Archaeoptéryx. [1] [2]




Références:
[1] Xu Xing et al (2011), An Archaeopteryx-like theropod from China and the origin of Avialae, Nature
[2] Lawrence M. Witmer (2011), An icon knocked from its perch, Nature
[3] Michael S. Y. Lee and Trevor H. Worthy, Likelihood reinstates Archaeopteryx as a primitive bird, The Royal Society
[4] Gerald Mayr et al (2005), A Well-Preserved Archaeopteryx Specimen with Theropod Features, Science
[5] Xu Xing et al (2010), Pre-Archaeopteryx coelurosaurian dinosaurs and their implications for understanding avian origins, Chinese Science Bulletin
En effet avant la découverte de Xiaotingia zhengi, Archaeoptéryx a généralement été considéré comme étant le «premier oiseau», c’est-à-dire le représentant le plus basal de la gente avienne. Concrètement cela signifie qu’Archaeoptéryx était phylogénétiquement plus apparenté aux oiseaux (modernes ou disparus) qu’aux autres dinosaures théropodes non-aviens, tels que le très célèbre Vélociraptor (voir arbre phylogénétique ci-dessous).

Mais donc la découverte de Xiaotingia zhengi remettrait en question cette plus grande proximité d’Archaeoptéryx avec les oiseaux jusque-là admise par la majorité des spécialistes. En effet les auteurs ayant analysé et décrit en détail le fossile de Xiaotingia zhengi en arrivèrent à la conclusion, que Archaeoptéryx serait davantage apparenté à aux dinosaures théropodes Deinonychosauriens comme Vélociraptor, qu’aux oiseaux moderne (voir arbre phylogénétique ci-dessous). [1] [2]

Tout cela a fait pas mal de bruit chez les spécialistes même si l’idée qu’Archaeoptéryx ne soient pas davantage apparenté aux oiseaux que ne l’est Vélociraptor, avait déjà été proposé par certains chercheurs des années avant la découverte de Xiaotingia zhengi.
Cependant les auteurs de l’étude du fossile de Xiaotingia zhengi, avaient pris soin de préciser que cette «exclusion» d’Archaeoptéryx du clade des oiseaux, n’était pas certaine et était susceptible d’être remise en question. Or par plus tard que ce mois d’octobre 2011, une nouvelle étude cladistique portant sur Archaeoptéyx et d’autres Dinosaures Théropodes, replace Archaeoptéryx à la base de la lignée avienne. (Voir schéma ci-dessous) [3]

Archaeoptéryx en rouge est à nouveau placé à la base de la lignée avienne, Notez également que selon cette étude Xiaotingia zhengi également indiqué ici en rouge, est placé à la base de la lignée de Troodontidés. [3]
Mais alors comment s’y retrouver? Un coup Archaeoptéryx est étroitement associé à la lignée avienne, une autre fois il en est exclus puis il y est à nouveau remis.
La seule réponse objective que l’on peut donner à cette problématique est que la position phylogénétique exacte d’Archaeoptéryx demeure incertaine.
Mais cependant ne nous y trompons pas non plus la position phylogénétique générale d’Archaeoptéryx, nous savons qu’Archaeoptéryx est un Dinosaures Théropode, Cœlurosaurien, Maniraptorien et qui plus membre du clade des Paraves (les Paraves étant le clade regroupant les Deinonychosauriens et les Avialae comprenant notamment les oiseaux).

Le clade des Paraves correspond à l'embranchement 14 du présent cladogramme. Notez qu'il existe des phylogénies alternatives, celles-ci ne concernant d'ailleurs pas que Archaeoptéryx mais également d'autres lignées. Ainsi par exemple Gregory Scott Paul proposa une phylogénie bouleversant totalement le clade des Paraves, puisque selon lui les Oviraptorosauria (embranchement numéro 11 du présent cladogramme) pourrait-être plus apparenté au clade des avialae (embranchement 15) que ne le sont les Deinonychosauriens (embranchement 18). Cependant même si cette phylogénie alternative est plausible et ne peut d'être exclue, la phylogénie ci-dessus demeure la plus soutenue par les analyses cladistique et donc la plus communément admise à ce jour.
C’est au sein du dit clade des Paraves que les choses se compliquent, car Archaeoptéryx semble être un proche cousin de l’ancêtre commun de la lignée ayant mené au Deinonychosauriens comme Vélociraptor et à celle ayant vu naître les oiseaux. Cette proximité probable avec l’ancêtre commun en question fait que le positionnement d’Archaeoptéryx au sein du clade des Paraves est sujet à caution, surtout qu’il existe encore deux à trois autres phylogénies alternatives pour le seul clade des dits Paraves! [4] [5]
En conclusion nous pouvons donc dire que Archaeoptéryx ainsi que les divers autres dinosaures à plumes retrouvés à ce jour, nous montrent à quel point l’évolution des Dinosaures Théropodes, Cœlurosauriens, Maniraptoriens a été buissonnante. De nombreuses lignées de dinosaures à plumes plus ou moins petits sont apparues et ont continué à évolué de leur côté de la fin du Jurassique et durant tout le Crétacé. Parmi ces divers lignées de dinosaures à plumes, l’une s’est grandement spécialisée dans le vol battu, il s’agit bien évidemment de celle des oiseaux.
Il reste maintenant à espérer que de nouvelles découvertes fossiles puisse un jour permettre d’affiner la positons de chacun de ces petits dinosaures à plumes Maniraptoriens, Archaeoptéryx compris, et ainsi permettre de mieux encore davantage en lumière l’origine évolutive de nos amis à plumes.
Références:
[1] Xu Xing et al (2011), An Archaeopteryx-like theropod from China and the origin of Avialae, Nature
[2] Lawrence M. Witmer (2011), An icon knocked from its perch, Nature
[3] Michael S. Y. Lee and Trevor H. Worthy, Likelihood reinstates Archaeopteryx as a primitive bird, The Royal Society
[4] Gerald Mayr et al (2005), A Well-Preserved Archaeopteryx Specimen with Theropod Features, Science
[5] Xu Xing et al (2010), Pre-Archaeopteryx coelurosaurian dinosaurs and their implications for understanding avian origins, Chinese Science Bulletin
dimanche 30 octobre 2011
Excréments racialistes, fin.....(provisoire).....

Suite à la publication de mon message initiale sur le racisme «scientifique», un troll racialiste répondant au pseudo de darwincwg, mythomane se prétendant «docteur en biologie» (rien que cela) s'est donc sans surprise sentit obligé de me répondre avec virulence (au point de souhaiter ma mort) à plusieurs reprises. Ses réponses consistait notamment en de longs copiés/collés issu de sites racialistes. Lui ayant refuser ce flood (car la fonction «commentaires» est réservé aux commentaires et pas au flood), il m'a donc répondu tant bien que mal avec ces mots à lui, même si il se contentait donc toujours de pomper mécaniquement ce qui se trouvait sur ses sites racialistes de référence avec notamment comme référence de base d'illustres racialistes tel que John Philippe Rushton (voir photo ci-dessus).....
Comme on pouvait s'y attendre ses réponses étaient donc insipides, car ignorant les objections déjà amené à ces «arguments», et bien évidemment ignorant les nouvelles objection que je lui amenait, références à l'appui, et donc se contentant de répéter en boucle les mêmes sophismes. Mais le pire étant son absence totale de maîtrise en matière de biologie, et plus spécifiquement en matière de génétique et d'anthropologie. Son incapacité à argumenter l'ayant au final poussé, à souhaiter ma mort et autres joyeusetés pour finalement l'amené à aller me répondre sur son blog à lui à coup de longs copiés/collés ignorant bien évidemment tout ce qui lui avait été expliqué.
Jugeant à juste titre inutile de répondre à un troll ne maîtrisant strictement rien aux problématiques scientifiques qu'il aborde et ignorant tout ce qu'on lui explique en plus de distordre les propos de ses contradicteur, je n'avait initialement pas pensé à rédigé un nouveau message ne guise de réponse. Néanmoins deux de ces affirmations fallacieuse en matière de biologie, notamment de génétique, me semble intéressant à corriger non pas dans l'espoir de refaire l'éducation perdue et/ou inexistante du troll en question, mais simplement pour exposer clairement le caractère fallacieux des «démonstrations» racialistes. Ce message étant, je l'espère, le dernier consacré aux inepties racialistes, avant longtemps.
__________________________________________
Nous pouvons revenir sur les propos de ce docteur en biologie auto-proclamé, ne maîtrisant même pas le quart du tiers de la discipline dont il se dit spécialiste. Mais avant cela souvenons que darwincwg avait affirmé que le noirs africains seraient génétiquement plus proche d'Homo erectus que les européens et les asiatiques. Je lui avait demandé de me fournir une source démontrant cela. Et en guise de référence darwincwg se référa donc à ce papier de John Philippe Rushton, papier qui n'amène strictement aucune donnée génétique! En effet pour darwincwg, pas besoin de données génétique pour démontré que les africains seraient génétiquement plus proche d'Homo erectus car John Philippe Rushton l'aurait démontré via des variables phénotypiques. À ce titre voilà comment darwincwg expose la chose.

J’ignore comment on peut être suffisamment à la masse pour se prétendre «docteur en biologie» et affirmer une pareille énormité..... Ah oui c’est vrai il suffit d’être complètement à la masse bien sûr!
Il y a en effet plusieurs choses que le mythomane darwincwg ignore en matière de biologie mais également plus spécifiquement en matière de paléoanthropologie.
1. Des traits similaires peuvent-être apparus indépendamment dans deux ou plusieurs populations. Par exemple une étude relativement récente sur certaines séquences d’ADN néanderthaliennes suggèrent, que certains néanderthaliens étaient probablement roux comme certains européens modernes. Pourtant les variations génétiques qui auraient confié cette « rousseur» à certains néanderthaliens, ne sont pas les mêmes que celles des européens roux d’aujourd’hui. [1] Bref un caractère phénotypique similaire voir identique, n’a pas forcément une origine génétique commune et ne témoigne pas forcément d’un lien de parenté.
2. Des caractères similaires à certaines populations d’Homo erectus ont été retrouvé chez les populations…..Est-Asiatiques. Étrange sachant car selon les standards de Rushton (et donc de darwincwg) les Est-Asiatiques devraient être plus «éloignés» des Homo erectus. Les caractères Est-Asiatiques en question étant notamment les incisives en pelle + d’autres traits au niveau du crâne. Certains paléoanthropologues suggérant un métissage entre les Homo erectus asiatiques et les Homo sapiens sapiens venus d’Afrique [2]. D’autres penchent pour une simple convergence évolutive.
3. Si l’on veut prouver que les noirs-africains sont génétiquement plus proches d’Homo erectus que les autres populations humaines, il faudrait retrouver de l’ADN d’Homo erectus et si possible de différentes populations d’Homo erectus à travers le monde (car Homo erectus était répandu de l’Afrique à l’Est de l’Eurasie avec des variations notables d’une population à l’autre) de comparer tout cela aux données génétique existante des populations humaines actuelles, pour être fixé.
Mais la faute la plus grave de Darwincwg demeure sa vision même de l’évolution en général et de l’évolution humaine en particulier. En effet parler «d’échelle de l’évolution» est totalement obsolète de nos jours. Cette vision «gradiste» de l’évolution où l’on placerait donc Homo erectus en amont et les africains entre Homo erectus et les eurasiens, est donc une stupidité crasse ignorant aussi bien les modalités réelles de l’évolution en général que la complexité de l’évolution humaine récente en particulier.
C’est d’ailleurs aussi en cela que le papier de Rushton auquel il se réfère, n’amène aucune donnée génétique et n’amène plus généralement strictement aucune démonstration rigoureuse des thèses racialistes qu’il défend.
Rushton ne faisant dans son papier que deux choses- à savoir tout d’abord enfoncer des portes ouvertes pour ensuite prétendre que ces thèses racialistes seraient donc elles aussi des évidences avérées.
Premièrement Rushton ne fait que constater via des corrélations ce que toute personne un minimum informer en matière d’évolution humaine sait déjà. À savoir que la largeur du bassin et volume crâniens on co-évolué, chez les hominidés, une tête plus grosse nécessitant un bassin plus large pour permettre la parturition. Et bien sûr un bassin plus large ayant quelques peu modifié les insertions fémorales avec une incurvation fémorale plus grande.
Fort d’avoir prouvé que la Terre est ronde plus de 500 ans après Christophe Colomb, Rushton ajoute alors que l’on constaterait un moindre volume crânien, une moindre largeur du bassin, et une moindre incurvation fémorale chez les africains actuels que chez les eurasiens. Tout cela étant sensé confirmer ses thèses racialistes selon lesquels les africains seraient non-seulement moins intelligent mais également auraient moins «évolué» depuis Homo erectus que ne l’ont fait les populations eurasiennes!
Rien que cela!
Mais il y a bien évidemment plusieurs problèmes qui sont totalement ignoré par cette pseudo-étude de Rushton.
1. nombreuses données absentes ou pire encore une absence d’informations sur certaines données.
2. Concernant les données des populations humaines actuels, il n’y aucune prise en compte des variations existant au sein des grand regroupements continentaux.
3. Cette pseudo-étude ignore des études et observations antérieures ayant montré en quoi les variations existant au sein des grands regroupements continentaux. rendent ces derniers non-pertinents pour déterminer des «tendances» évolutives, comme cela est fait entre les différentes espèces d’hominidés!
Par exemple Rushton s’était donné au même raisonnement avec la maturation. En effet l’analyse du squelette d’un jeune Homo erectus laisse penser qu’Homo erectus avait une maturation bien plus rapide que la nôtre. En fait il semblerait même que la maturation d’Homo erectus soit plus similaire à celle des grands singes qu’à celle d’Homo sapiens sapiens. En cela les auteurs de cette étude parue dans la revue Nature pensent qu’Homo erectus avait probablement des capacités cognitives nettement moindres que celles des hommes modernes. [3] Or là encore John Philippe Rushton prétend que les noirs-africains ont une maturation plus rapide que les autres populations humaines en se basant là encore sur quelques données agrégées.
Ainsi la plus grande maturation ferait à la fois office de «démonstration raciale» tout en confirmant les idées de Rushton voulant que les populations noires africaines aient moins évolués en matière de maturation et d’intelligence, depuis Homo erectus.
Bien évidemment les conclusions de Rushton sont bancales car ignorant s’adonnant là encore à des agrégations ignorant la diversité génétique et phénotypique humaine dans son ensemble, en plus de prendre parti gratuitement pour une vision simpliste et linéaire de l’évolution humaine. Premièrement les différences de rythme de maturation entre les différentes populations humaines sont nettement moindre que celle estimée entre Homo erectus et l’homme moderne. De plus les différences moyennes de maturation entre populations humaines ne se répartissent pas de manière géographiquement nettes comme semble le suggérer Rushton et ne permettent pas de déduire quoi que ce soit en ce qui concerne l’intelligence des populations humaines en question. Tout cela n’étant pas surprenant sachant que l’évolution humaine n’est guère aussi linéaire que semble le percevoir Rushton.
Par exemple en 1974 le psychologue Daniel G. Freedman constata qu’une ethnie «caucasienne» du nord de l’Inde les Punjabi, avaient une maturation très rapide en comparaison des autres populations. En fait la maturation des enfants de cette ethnie, était alors la plus rapide connu à travers le monde, dépassant les moyennes calculé pour les populations noires africaines. Notons également cette ethnie, loin de confirmer les thèses de Rushton mettant en corrélation maturation, intelligence et succès économique, faisait preuve d’un certain succès économique.
L’anthropologue Léonard Liebermann est revenu sur les enseignement des observation de Freedman [4]
Premièrement la rapidité de maturation varie énormément au sein des «races». Il est ainsi difficile d’affirmer que les moyennes faites entre macro-populations (noirs-africains, caucasiens, asiatiques) soient significatives et donc trahissent quoi que ce soit en matière de proximité phylogénétique.
Deuxièmement, les variations inter-populations dans la rapidité de la maturation semblent être indépendantes de la «race» et des aptitudes intellectuelles des populations en question. Elles semblent trahir des évolutions locales (au sein des grands regroupements appelés «races»), peut-être en raison de pressions sélectives particulières, mais donc sans que cela puisse être rattaché à des aptitudes intellectuelles particulières.
Tout cela est bien sûr ignoré par Rushton et compagnie, ces derniers ne jugeant que par des agrégations dont la pertinence n’est pas démontré voir même sérieusement mise à mal, ainsi que par des déductions spéculatives ayant pour base une vision linéaire, «gradiste» et donc fausse de l’évolution.
Enfin on peut également noter un point amusant. Des études ont montré que l’Homme de Néanderthal avait lui aussi une maturation plus rapide que la nôtre [5] [6] et plus récemment une étude a montré que les eurasiens étaient de toute évidence génétiquement plus proche de Néanderthal que ne le sont les africains (car les ancêtres des eurasiens se seraient métissé dans une certaine mesure avec des Néanderthaliens). Question darwincwg pourra-t-il intégré cet exemple pour comprendre pourquoi on ne peut pas inférer de manière simple une supposée proximité génétique à partir de caractéristiques phénotypiques?
Bref l’agrégation de macro-populations continentales faite par Rushton est fallacieuse. Mais pire pareille agrégation tient d’autant moins avec les populations noires africaines lorsque l’on sait que celles-ci sont très diverses les unes par-apport aux autres et que l’ensemble eurasien n’est en réalité qu’un sous-ensemble d’une diversité génétique essentiellement noire africaine. [7]

Comme le montre cet «arbre d'apparentement» ci-dessus l’analyse du génome de divers populations africaines et non-africaines, montrent la très grande diversité génétique africaine en comparaison avec celle du reste du monde qui n’en constitue qu’en fait un sous-ensemble. Aucune des agrégation raciales de Rushton consistant à présenter les noirs-africains comme un ensemble cohérent, ne tient compte de cette complexité et notamment des relations phylogénétique des diverses populations noires-africaines. Pourtant cette complexité peut rendre les moyennes entre populations continentales de Rushton totalement caduque. Mais Rushton ne s’embarasse pas de ces données, ignore leurs implications et préfère donc s’en tenir mordicus à ses agrégations bancales en guise de démonstration.
Et donc il ne suffit pas de sortir une moyenne des « noirs africains » pour telle ou telle mesure phénotypique, il faudrait plutôt établir des moyennes des diverses populations africaines, en tenant compte des divergences et proximités phylogénétique des diverses populations noires-africaines en question. Et il faudrait s’assurer que l’échantillonnage des différentes populations, est donc pertinent compte tenue de ces proximités phylogénétiques.
À ce titre prenons un exemple concret concernant les traits musculo-squelettiques.
Le grand médecin, anatomiste et anthropologue français du 19ème siècle Paul Broca était, comme la plupart des hommes occidentaux de classe sociale supérieure de l'époque, convaincu de l’infériorité intellectuelle des noirs, qu’il percevait comme étant plus «simiens» que les blancs. Un des caractères musculo-squelettique que Paul Broca mesura était le rapport radius (os de l’avant-bras) – humérus (os du bras à proprement parler). Selon Broca plus l’avant-bras est long par apport aux bras plus cela se rapproche d’une condition «simienne». Or Broca était content de constater que les noirs africains avaient en moyenne un avant-bras plus long par-apport aux bras, comparé aux européens. Cependant les choses se compliquèrent lorsqu’il mesura ce rapport de longueur chez des représentant de l’ethnie khoïkhoï, pour qui l’avant-bras était bien moins long par apport à l’avant-bras que cela n’était le cas chez les blancs! [8] Bref là encore nous avons une donnée exposant le caractère fallacieux des agrégations faites par John Philippe Rushton!
Car ce que fait Rushton en conglomérant les noirs africains, sans même que les échantillonnages prennent en compte cette diversité des dits noirs-africains, revient exactement au même que de mesurer la taille moyenne des noirs-africains versus la taille moyenne des blancs européens. Que signifierait la taille moyenne des noirs africains alors que l’on trouve un vaste gamme des variations chez ces derniers avec des pygmées de taille très inférieure à la moyenne et d’autres populations africaines ayant au contraire une taille moyenne remarquablement grande? D’ailleurs comment réaliser un échantillonnage pertinent pour déterminer la taille moyenne des noirs africains compte tenus de ces variations importantes et complexes dans leurs répartitions respectives, d’une population noire africaine à l’autre? Car si l’on surreprésente les pygmées ou inversement notre moyenne sera forcément biaisée et donc non-pertinente. La même question se poserait d’ailleurs en grande partie pour les européens pourtant moins divers sur le plan génétique.
Mais John Philippe Rushton ignore bien sûr ce dernier point, la rigueur méthodologique ne l’intéresse pas, seule la validation même bancale mais emballé d’une belle rhétorique de ses préjugés, comptent.
En conclusion il n’existe aucune démonstration génétique. Et la soit disant démonstration phénotypique n’a même pas un quart de tiers de rigueur nécessaire pour permettre d'extrapoler des tendances évolutives inter-espèce à une diversité phénotypique intra-espèce. Normal puisque John Philippe Rushton ignore, comme à son habitude, la complexité de la diversité humaine et ne jure que par l’agrégat de quelques variables en ignorant totalement la faiblesse de pareille démarche.
Mais avant de conclure définitivement ce message je reviens sur une dernière perle de darwincwg.

Non, il n’est pas correct de dire que l’homme descend du singe, car cela prête à confusion sachant qu’en réalité l’homme est lui-même biologiquement parlant, un singe! L’espèce humaine est une espèce de singe parmi d’autres (bien que donc dotés de particularités remarquables), une espèce de singe qui partage des ancêtres communs avec les autres singes. Les ancêtres communs en question étaient bien évidemment également des singes, mais donc ces ancêtres communs n’existent plus aujourd’hui.
Ainsi si l’on prend nos plus proches cousins actuels que sont les bonobos et les chimpanzés, nous partageons avec des derniers un ancêtre commun dont l’âge est estimé entre 5 et 10 millions d’années (probablement aux alentours de 6 à 8 millions d’années). Nous descendons donc de cet ancêtre commun mais pas du chimpanzé. L’affirmation «l’homme descend du singe» prêtant à confusion, n’exprimant pas pertinemment l’évolution humaine telle qu’elle a eu lieu et donc n’étant guère correct.
Tout cela nous amène à l’affirmation de darwincwg selon laquelle l’on pourrait dire que les européens descendent des africains et que cela serait là aussi correct.
Or ce n'est bien évidemment pas le cas!
D’une part parce que là encore l’on raisonne en terme d’ancêtres communs. Mais aussi et surtout parce que contrairement à la lignée du chimpanzé avec laquelle nous n’avons plus de flux de gènes depuis plusieurs millions d’années, il y a eu de flux de gènes conséquent entre l’Europe et l’Afrique il y a encore moins de 40'000 ans!
Darwincwg cite le généticien Luigi Luca Cavalli-Sforza pour affirmer par exemple que les africains seraient apparus il y 200'000 ans et les européens il y 100'000 ans. Mais il y a de nombreuses inexactitudes dans cette affirmation. La question serait de savoir si Cavalli-Sforza a réellement fait une erreur, s’est mal exprimé ou que Darwincwg l’a mal compris et/ou retranscrit. Toujours est-il que l’on peut corriger cette affirmation en rappelant les trois choses suivantes.
1. L’apparition de l’homme moderne (Homo sapiens), est estimé avoir eu lieu il y a 200'000 ans en Afrique.
2. Si des sorties hors d’Afrique de la part de l’homme moderne ont probablement déjà eu lieu il y plus de 100'000 ans, il semble qu’elles contribuèrent que de manière limité au patrimoine génétique de l’humanité moderne. Par ailleurs à ce jour rien ne prouve qu’il y a 100'000 ans l’homme moderne avait colonisé l’Europe qui semblait alors encore peuplée exclusivement de Néanderthaliens. Par ailleurs le chromosome Y indique des sorties du continent africain largement plus récentes qu'il y a 100'000 ans! [9]
3. L’homme moderne est arrivé en Europe aux alentours de 50'000 à 40'000, où hormis les premiers fossiles européens d’Homo sapiens l’on observe également les premiers artefacts culturels associés à Homo sapiens avec entre autre l’implantation de la «Culture Aurignacienne». [10]
À tous ces éléments mettant déjà à mal l’affirmation de darwincwg, il est nécessaire de rappeler à nouveau que les données génétiques actuels indiquent l’existence de flux de gènes conséquents allant de l’Afrique subsahariene à l’Europe il y moins de 40'000 ans voir même probablement il y a 20'000 ans seulement. [11]
D’ailleurs l’existence de pareilles flux de gènes relativement récent dans la préhistoire humaine se remarque aisément lorsque l’on observe la distribution de l’haplogroupe T du chromosome Y!

L’haplogroupe T du chromosome Y a une distribution allant de l’Afrique Subsaharienne, à l’Europe et même jusqu’en Inde. La date d’apparition de cet haplogroupe se situe entre 19'000 et 34'000 ans et trahit donc des flux de gènes relativement récent entre différents continents notamment entre l’Europe et l’Afrique. [12]
Bref les européens ne descendent pas des africains ils partagent divers ancêtres communs récents, voir même étonnamment récents, qui vivaient selon toute vraisemblance en Afrique. Et mieux encore divers flux de gènes ont continué d’avoir lieu entre les populations africaines et européennes à des temps bien plus récents encore.
Ces mises au point étant faite je pense pouvoir fermer provisoirement cette petite série de message sur le racisme «scientifique». Tout ce que l’on peut retirer de l’histoire est que le racialisme, c’est-à-dire ces doctrines raciales visant à dépeindre certaines populations comme «inférieures» ou «nuisibles» en raison de leur «biologie», a pour seules bases la distorsion de données anthropologiques, l’absence de rigueur et une indécrottable mauvaise foi! Et il est bien sûr aussi illusoire d’essayer de convaincre un fanatique décérébré racialiste qu’un créationnistes. Surtout lorsque le racialiste en question est suffisamment à la masse pour s’autoriser des assertions mythomanes aussi énorme que celles consistant à se faire passer pour un docteur en biologie sans même avoir ne serait-ce qu’un quart du tiers de bagage en la matière!
[1] Carles Lalueza-Fox et al (2007), A Melanocortin 1 Receptor Allele Suggests Varying Pigmentation Among Neanderthals, Science
[2] Hong Sang et al (2007), An early modern human from Tianyuan Cave, Zhoukoudian, China, Proceedings of the National Academy of Sciences
[3] Hélène Coqueugniot et al (2004), Early brain growth in Homo erectus and implications for cognitive ability, Nature
[4] Leonard Lieberman (2001), How “Caucasoids” Got Such Big Crania and Why They Shrank, Current Anthropology, From Morton to Rushton
[5] Tanya M. Smith et al (2007), Rapid dental development in a Middle Paleolithic Belgian Neanderthal, Proceedings of the National Academy of Sciences
[6] Marcia S. Ponce de Leoet al (2008), Neanderthal brain size at birth provides insights into the evolution of human life history, Proceedings of the National Academy of Sciences
[7] Sarah A. Tishkoff et al (2009), The Genetic Structure and History of Africans and African Americans, Science
[8] Paul Broca (1862), Sur les proportions relatives du bras, de l’avant-bras et de la clavicule chez les noirs et les Européens, Bulletins Société d’Anthropologie, Paris 3: 32pp
[9] Peter A. Underhillet al (2000), Y chromosome sequence variation and the history of human populations, Nature
[10] Michael Bolus et Nicholas Conard (2011), Les débuts de la culture en Europe, Pour La Science, numéro 407 -Septembre 2011
[11] Heng Li and Richard Durbin (2011), Inference of human population history from individual whole-genome sequences, Nature
[12] Fernando L. Mendez et al (2011), Increased Resolution of Y Chromosome Haplogroup T Defines Relationships among Populations of the Near East, Europe, and Africa, Human Biology
lundi 26 septembre 2011
Pseudoscience racialiste, suite.....

Nouvelle réponse adressée au racialiste répondant au pseudo de darwincwg et qui ne tient toujours pas compte, ni ne comprend les explications qui lui sont fournies.
___________________________________________________________
Darwincwg, je dois dire que ta mauvaise foi est pour le moins déplorable. Tu ignores les arguments et sources qui te sont adressé pour répéter en boucle les mêmes sophismes dont il a déjà été démontré qu'il s'agit.....de sophismes. De plus dans un de tes commentaires tu as d’abord voulu m’assimiler de façon parfaitement gratuite et mensongère, à un gus qui soutiendrait l’idée d’une hérédité nulle concernant les différences «d’intelligence» entre individus. Lorsque je te rappelle alors que je n’ai jamais nié le fait qu’il existe bien évidemment une influence génétique en matière de psyché entre les individus, tu me réponds ceci.
Darwincwg, as-tu simplement ne serait-ce que le quart du tiers de notions élémentaires en matière de génétique des populations, d’hérédité et plus spécifiquement de paléoanthropologie?
Tout d’abord on parle de fréquences alléliques, les allèles étant différentes version de mêmes gènes. Deuxièmement il n’y a pas deux individus (à l’exception des vrais jumeaux) qui ont le même génotype. Et si l’on raisonne en termes de populations tu trouves des différences de fréquences alléliques entre l’Europe du Nord et du Sud, de l’Est et de l’Ouest ou même entre deux pays voisins. Bref les différences de fréquences alléliques ne correspondent pour la plupart pas aux grandes « subdivisions raciales » comme on les appelle parfois, ces dernières étant des virtualités statistiques basées sur des corrélations entre les différentes fréquences de différents allèles de certains gènes (voir par exemple les études utilisant l’indice Fst souvent utilisé en la matière). [1]
Enfin non les africains et les européens n’ont pas vécu séparé depuis 100'000 ans…..Car par exemple le dernier ancêtre masculin commun de l’humanité moderne nous ayant transmis son chromosome Y (le fameux «Adam Chromosomique»), est estimé a bien moins de 100'000 ans. [2] Mais surtout une récente étude montre l’existence de probables flux gènes ayant eu cours entre l’Eurasie et l’Afrique aux alentours d’il y a 40'000 à 20'000 ans seulement! [3]
Dois-je te rappeler que les divergences génétiques entre les populations modernes à travers le monde, malgré les différences en termes de fréquences alléliques existantes, sont bien moindre que les divergences génétiques existantes entre les différentes populations de chimpanzés? [4] [5] [6]

___________________________________________________________
Darwincwg, je dois dire que ta mauvaise foi est pour le moins déplorable. Tu ignores les arguments et sources qui te sont adressé pour répéter en boucle les mêmes sophismes dont il a déjà été démontré qu'il s'agit.....de sophismes. De plus dans un de tes commentaires tu as d’abord voulu m’assimiler de façon parfaitement gratuite et mensongère, à un gus qui soutiendrait l’idée d’une hérédité nulle concernant les différences «d’intelligence» entre individus. Lorsque je te rappelle alors que je n’ai jamais nié le fait qu’il existe bien évidemment une influence génétique en matière de psyché entre les individus, tu me réponds ceci.
Aaah... les gènes sont donc responsables des différences intra-raciales observées... mais pas interraciales ? Les africains et les européens, qui ont vécu dans des milieux séparés pendant 100 mille ans, ont les même génotypes intellectuels, en terme qualitatif et de fréquence de gènes ? C'est très vraisemblable dites-moi tout cela, puissamment raisonné. darwincwg
Darwincwg, as-tu simplement ne serait-ce que le quart du tiers de notions élémentaires en matière de génétique des populations, d’hérédité et plus spécifiquement de paléoanthropologie?
Tout d’abord on parle de fréquences alléliques, les allèles étant différentes version de mêmes gènes. Deuxièmement il n’y a pas deux individus (à l’exception des vrais jumeaux) qui ont le même génotype. Et si l’on raisonne en termes de populations tu trouves des différences de fréquences alléliques entre l’Europe du Nord et du Sud, de l’Est et de l’Ouest ou même entre deux pays voisins. Bref les différences de fréquences alléliques ne correspondent pour la plupart pas aux grandes « subdivisions raciales » comme on les appelle parfois, ces dernières étant des virtualités statistiques basées sur des corrélations entre les différentes fréquences de différents allèles de certains gènes (voir par exemple les études utilisant l’indice Fst souvent utilisé en la matière). [1]
Enfin non les africains et les européens n’ont pas vécu séparé depuis 100'000 ans…..Car par exemple le dernier ancêtre masculin commun de l’humanité moderne nous ayant transmis son chromosome Y (le fameux «Adam Chromosomique»), est estimé a bien moins de 100'000 ans. [2] Mais surtout une récente étude montre l’existence de probables flux gènes ayant eu cours entre l’Eurasie et l’Afrique aux alentours d’il y a 40'000 à 20'000 ans seulement! [3]
Dois-je te rappeler que les divergences génétiques entre les populations modernes à travers le monde, malgré les différences en termes de fréquences alléliques existantes, sont bien moindre que les divergences génétiques existantes entre les différentes populations de chimpanzés? [4] [5] [6]

Schéma illustrant les distances génétique mesuré avec l’ADN mitochondrial, entre les populations de différentes espèces. Malgré un échantillonnage plus vaste (811 individus pris sur différents continents), l’on trouve moins de distance génétique entre les différentes populations continentales humaines, qu’entre les différentes sous-populations de chimpanzés ou de gorille pourtant présentes sur le seul continent africain. Cela témoignant de la grande proximité génétique et phylogénétique des populations humaines actuelles. Ces résultats étant confirmé par des études portant sur le génome à proprement parler. Schéma tiré de Pascal Gagneux et al (1999) [7]
Ces mises au point étant faites il reste donc la question des différences «d’intelligence innée» supposées entre les différentes populations humaines actuelles. Chose sur laquelle je reviens plus en détail plus bas dans mon présent message.
Néanmoins je précise déjà que:
1. Il faudrait déjà disposer d’une définition claire de ce qu’est «l’intelligence» qui ne peut certainement pas se réduire aux seuls succès dans les réponses fournies à une série de tests standardisés à savoir les tests de QI.
2. Déterminer que les divergences de QI entre populations s’expliquent réellement de manière prépondérante par des différences de fréquences alléliques et donc contrôler via des protocoles expérimentaux rigoureux, les influences environnementales afin de les dissocier des facteurs génétiques.
Encore une fois j’aborderai plus en détail ces deux points plus bas, références à l’appui cela va de soi.
Mais tout d'abord revenons sur la suite de ta prose darwincwg.
Ta présente prose darwincwg prouve encore une fois tes profondes lacunes en matière de biologie de l’évolution et d’évolution humaine en particulier.
1. Les mutations n’apparaissent pas en fonction de l’environnement mais indépendamment de celui-ci, les mutations se produisant de manière aléatoire. Seule la sélection est susceptibles de favorisé telle ou telle mutation c’est-à-dire de sa fixation au sein d’une population. De plus la sélection n’est pas seule à entrer en jeu la dérive génétique a également une grande importance dans la fixation ou non de certains allèles et donc l’évolution des populations.
2. Rien ne prouve que les peuplades vivant plus au Sud n’ont pas eu des pressions sélectives moins moins importantes que les peuplades vivant dans les climats froids, et susceptibles de favoriser les individus les plus «intelligents» (quoi qu’il faudrait encore une fois déjà préciser ce que l’on entend exactement par-là). Les forêts tropicales comprennent divers dangers et sont propices à bon nombre de maladies De plus pendant la glaciation une bonne partie du continent africain connu une intense désertification avec à la clef raréfaction des ressources et donc nécessité de s’adapter.
Ces mises au point étant faites il reste donc la question des différences «d’intelligence innée» supposées entre les différentes populations humaines actuelles. Chose sur laquelle je reviens plus en détail plus bas dans mon présent message.
Néanmoins je précise déjà que:
1. Il faudrait déjà disposer d’une définition claire de ce qu’est «l’intelligence» qui ne peut certainement pas se réduire aux seuls succès dans les réponses fournies à une série de tests standardisés à savoir les tests de QI.
2. Déterminer que les divergences de QI entre populations s’expliquent réellement de manière prépondérante par des différences de fréquences alléliques et donc contrôler via des protocoles expérimentaux rigoureux, les influences environnementales afin de les dissocier des facteurs génétiques.
Encore une fois j’aborderai plus en détail ces deux points plus bas, références à l’appui cela va de soi.
Mais tout d'abord revenons sur la suite de ta prose darwincwg.
Les différences intellectuelles entre les races sont conséquentes des différents environnements dans lesquels elles ont vécu, avec en particulier l’impact du principal âge de glace (-28000 à -10000) dans l'hémisphère nord exerçant des pressions de sélection pour une plus grande intelligence pour survivre.
Il y eu apparition de mutations pour plus d’intelligence dans les populations nombreuses et soumises au stress du froid et augmentation progressive par sélection naturelle des fréquences des gènes pour une plus haute intelligence. darwincwg
Ta présente prose darwincwg prouve encore une fois tes profondes lacunes en matière de biologie de l’évolution et d’évolution humaine en particulier.
1. Les mutations n’apparaissent pas en fonction de l’environnement mais indépendamment de celui-ci, les mutations se produisant de manière aléatoire. Seule la sélection est susceptibles de favorisé telle ou telle mutation c’est-à-dire de sa fixation au sein d’une population. De plus la sélection n’est pas seule à entrer en jeu la dérive génétique a également une grande importance dans la fixation ou non de certains allèles et donc l’évolution des populations.
2. Rien ne prouve que les peuplades vivant plus au Sud n’ont pas eu des pressions sélectives moins moins importantes que les peuplades vivant dans les climats froids, et susceptibles de favoriser les individus les plus «intelligents» (quoi qu’il faudrait encore une fois déjà préciser ce que l’on entend exactement par-là). Les forêts tropicales comprennent divers dangers et sont propices à bon nombre de maladies De plus pendant la glaciation une bonne partie du continent africain connu une intense désertification avec à la clef raréfaction des ressources et donc nécessité de s’adapter.
3. Si le climat froid génère réellement une plus grande «intelligence» comment expliquer le cas de l’homme de Neandertal? Neandertal ayant disparu suite à l’arrivé de l’homme moderne en Europe!
On sait que les premiers hommes modernes arrivés en Europe venaient de latitudes plus méridionales. La morphologie des premiers européens modernes se caractérisait par exemple par des membres plus longs par apport au tronc que la moyenne des européens actuels (là où les populations tropicales conservent en moyenne ces proportions de type «tropicale»). [8] Pourtant ces hommes modernes venus de climats plus chauds portaient avec eux des innovations culturelles que n’avaient jamais accomplis les Néanderthaliens [9] malgré des dizaines de milliers d’années de présence en Europe c’est-à-dire dans un climat plus froid. Et pire encore Neandertal disparu peu à peu laissant la place à Homo sapiens sapiens. Étrange car selon la thèse de Richard Lynn (oui darwincwg tes sources je les connais) cela devrait être exactement le contraire, Néanderthal soumis au climat plus rude aurait dut surpasser intellectuellement, culturellement et donc techniquement les Homo sapiens issu de climats plus chauds, ce qui ne faut absolument pas le cas!
Mais Richard Lynn ne s’embarrasse bien évidemment pas de prendre ne compte les faiblesses de ses thèses rendant donc toutes ses déductions racialistes bancales!
Les différences de QI entre les races expliquent les différences dans la capacité à faire la transition néolithique de chasse-cueillette à l'agriculture sédentaire, la construction des premières civilisations et le développement des civilisations matures durant les deux mille dernières années. La position des environnementalistes qui prétendent que depuis son apparition il y a 200000 ans, les hommes, séparés par des barrières géographiques dans différentes parties du monde, ont évolué en une dizaine de races différentes avec des différences marquées dans la morphologie génétique, les groupes sanguins, et l'incidence des maladies génétiques et pourtant auraient les mêmes génotypes pour l’intelligence, est si improbable que ceux qui l'avance doivent être totalement ignorants des principes fondamentaux de la biologie évolutive ou avoir un agenda politique pour nier de cette manière l'importance de la race. Ou les deux. darwincwg
J’imagine que tu disposes des mesures de QI des peuplades du Néolithique pour affirmer une telle chose…..Ah non c’est juste tu te réfères bêtement aux thèses de Richard Lynn qui ne contiennent bien évidemment aucune donnée concrète en la matière en plus de partir du postulat que l’intelligence peut se mesurer à l’aide de tests standardisés ne prenant même pas en compte l’ensemble des manifestations de la psyché pouvant être rangées dans le label «intelligence». Concernant les innovations ayant eu cours durant le Néolithiques Richard Lynn ignore par ailleurs les divers facteurs géographiques et environnementaux susceptibles d’expliquer les différences dans le développement des société ainsi que le fait que des peuplades d’Afrique noires avaient également développé agricultures et métallurgie bien avant notre ère. [10]
Autre point important, les caractéristiques telles que les différences de fréquences en matière de groupes sanguins et de maladies génétiques, ne définissent pas des «races» sauf à considérer comme formant une «race» n’importe quelle population différant en matière de fréquences alléliques par apport à une autre….. Auquel car les basques devraient alors être considéré comme étant une «race»! Et pour les différences morphologiques ce n’est non plus pas forcément aussi évident que de s’en servir pour s’adonner à des classifications raciales.

Même les différences phénotypiques visibles peuvent induire en erreur en matière de classification raciale.
Le dernier point étant que ce que tu appelles l’«intelligence» n’est pas formellement défini. Le mot «intelligence» lui-même étant un terme générique comprenant en réalité diverses manifestations complexes de notre psyché! C’est d’ailleurs là que Richard Lynn et autres dogmaticiens du QI se fourvoient gravement. Ce qu’on appelle l’intelligence englobes divers capacités cognitives, qui individuellement pour certaines pourraient être considéré comme étant des phénotypes, mais qui même individuellement sont tributaire de multiples gènes, de facteurs environnementaux et mêmes souvent d’autres capacités cognitives. C’est ainsi par exemple que des chercheurs ont remarqué qu’en entrainant sa mémoire, l’on augmente son «intelligence fluide» alors que la mémoire est sensée être indépendante de cette dernière. [11] Par ailleurs je le répète pour que cela soit clair, le plus grave étant que les tests de QI ne prennent de loin pas en compte l’ensemble des aptitudes cognitives susceptibles d’être rangées sous le label «intelligence».
Cette non prise en compte de manifestations cognitives importantes par les tests de QI ainsi que la complexité de notre psyché, des divers gènes et interactions de gènes entrant en compte ainsi que la manière exacte dont l’environnement peut modifier la dite psyché, ne sont bien évidemment pas envisagés par Richard Lynn!
Or sans envisager ces points et sans vraiment démêler la complexité de la psyché, l’affirmation de Richard Lynn selon laquelle les différences de QI entre populations évoluant dans des environnements sensiblement différents, prouvent une différence innée en matière d’intelligence, entre ces même populations, demeure une affirmation parfaitement gratuite!
En fait la notion même d’«intelligence» n’étant pas formellement définie on ne peut guère la quantifier avec un test standardisé qui aurait une portée universelle sans possible biais d’ordre social ou si tu préfères «anthropologique».
Aujourd’hui l’on est certes capable d’identifier l’influence probable que peuvent avoir certains allèles individuels sur certaines capacités, et encore de manière limité. Récemment l’on a été capable de faire une corrélation entre divers SNPs et des résultats à des «tests d’intelligence» en limitant au mieux, mais pas entièrement, les biais environnementaux. Mais on ignore quels sont les influences respectives de ces divers SNPs et de leurs interactions sur la psyché.
Enfin concernant les différences moyennes en matière de QI entre des populations évoluant dans des environnements sociaux différents il n’existe pas de démonstration en génétique montrant que les différences de moyennes sont d’origine génétiques. Et pour cause il n’existe pas de moyens de contrôler les variables environnementales pour les exclure et isolés les influences génétiques! Et le fait que la psyché est très malléable au contexte environnemental, c’est-à-dire social, éducatif et familial, rend pareil contrôle quasiment impossible.
J’avais déjà eu l’occasion dans ce message, d’expliquer comment en biologie on procède pour isoler les variables environnementales influant sur une population d’organismes, afin de déterminer l’influence du génome dans des variations phénotypiques très sensibles aux fluctuations environnementales (c’est-à-dire pouvant variés de façon importante en fonction de l’environnement, on parle alors d’une forte plasticité phénotypique). Pour cela on contrôle de manière stricte l’environnement de l’organisme et hormis l’échantillonnage qui se doit d’être important, l’on réitère si possible l’expérience avec plusieurs générations d’organismes toujours dans un environnement strictement contrôlé. [12]
Or aucune démonstration de cette rigueur n’existe dans la littérature «racialistes», les enfants noirs adoptés par les blancs ne respectant pas les critères de rigueurs requis car laissant paraître plusieurs biais potentiels. Et auxquels on peut opposer certaines données contradictoires. [13] Et le tout en ignorant le fait que bon nombre de psychologues mettent en garde contre l’usage non-contextualisé des tests de QI qui ne constitue pas un bête échelonnage du niveau d’intelligence quoi qu’en dise les Richard Lynn et compagnie. [14] [15]
Ainsi avec «l’intelligence» on se retrouve avec un problème non-résolu bien que l’on puisse cependant déjà déterminer quelques points intéressants.
Car par exemple même en admettant des différences de fréquences alléliques sensibles entre populations concernant des allèles impliquant des différences innées pour telle ou telle capacité cognitives, à quel point ces différences peuvent-elles exacerbées et/ou effacées par les facteurs environnementaux affectant également les capacités en question?
Pour répondre reprenons l’exemple d’un milieu institutionnalisé on constate que les enfants noirs ont en moyenne un QI plus élevé que les blancs [16] je pourrais en déduire que génétiquement les enfants noirs ont un avantage pour les capacités cognitives mesurés par les tests de QI! À cela tu me répondras sans doute que plein d’autres données statistiques montrent le contraire. Mais si aucune de tes données n’ont été faites en maîtrisant de manière stricte les variables environnementales, alors tu n’as guère de démonstration montrant que les enfants noirs ont un désavantage génétique, tout au plus mets-tu en doute mon affirmation selon laquelle les noirs auraient un avantage. Car tout ce que l’on est donc parvenu à montrer c’est que l’influence de l’environnement sur la moyenne des populations, détermine de manière très importante la moyenne de QI des populations! Elle l’influence bien plus que le font d’éventuelles différences en matière de fréquence d’allèles impliqués dans certaines capacités cognitives mesurées par le QI! À ce titre fie toi à ce sympathique article de Ned Block [15] qui devrait t’éclairer plus en détail sur la question de l’héritabilité! À ce titre voici un extrait de l'article en question qui devrait te faire réfléchir.
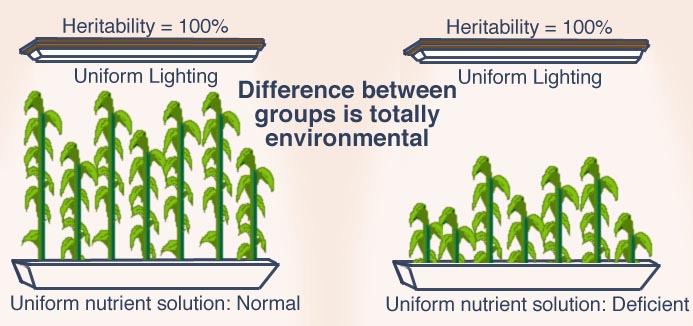
{kind=link}
{kind=link}
Supposez que vous achetiez un sachet de graines de maïs ordinaire. Vous en faites pousser une poignée dans un environnement bien contrôlé, où toutes les graines reçoivent la même lumière et la même solution nutritive. Les plants de maïs vont varier en hauteur. Comme l'environnement est le même pour tous, l'héritabilité - telle que définie plus haut - de la taille des plants sera de 100 %. Prenez maintenant une poignée du même sachet, et faites pousser les graines dans un environnement tout aussi uniforme, mais cette fois avec une solution nutritive insuffisante. Vous aurez à nouveau une variation de la taille de vos plants, mais tous seront rabougris. L'héritabilité de la hauteur des plants de maïs dans ce deuxième groupe est à nouveau de 100 %. Malgré une héritabilité de 100 % dans chaque groupe, la différence de taille entre les deux groupes est due entièrement à l'environnement. Ned Block [15]
Tous ces rappels darwincwg devant te permettre à présent de comprendre pourquoi, même en admettant l'importance de facteur génétiques dans les différences de QI d’un individu à l’autre, cela n’exclue de loin pas que les différence inter-population aient des causalités environnementales! Et cela d’autant plus que l’étude publié récemment dans la revue «Nature» corrélant les différences individuelles dans les résultats de QI et certains SNPs, n’arrive, pour les différences entre individus, qu'à une estimation d’environ 50% pour la contribution des polymorphismes génétiques. Et cela encore une fois en se limitant qui plus est aux seuls tests de QI, c’est-à-dire sans prendre en compte l’ensemble des manifestations de la psyché et de leurs interactions composant ce qu’on appelle communément «l’intelligence».
Ces rappels en matière de biologie, de méthodologie, d’hérédité et de psyché devraient te pousser à t’interroger sérieusement sur la crédibilité de Richard Lynn et des autres racialistes. Car considérer le QI comme un banal phénotype, affirmer pouvoir mesurer «l’intelligence» avec un seul chiffre sur une échelle unique tout en n’amenant aucune démonstration génétique sérieuse soutenant que les différences moyenne de QI entre populations sont d’origine génétiques, et étendre ces spéculations à la préhistoire humaine où les données et démonstrations sont encore moindres, c’est tout simplement de la charlatanerie!
Et cela devrait également te pousser à remettre sérieusement en question tes accusations à l’encontre de ceux qui rejettent les thèses racialistes pour ce qu’elles sont à savoir de la merde en barre!. Tu accuses en effet tes contradicteurs d’avoir un agenda idéologique et politique, sans même te demander si tes sources (Rushton, Jensen, Lynn et compagnie) ne pourraient pas elles aussi avoir le leur…..C’est amusant lorsque l’on sait que, par exemple que John Philippe Rushton est à la tête d’une organisation fondé par Wickliffe Preston Draper, dont les sympathies pour le régime nazi ne sont guère un secret et dont l’organisation avait clairement des objectifs idéologiques racistes et eugénistes…..Étrange car aujourd’hui cette même organisation est toujours centrée sur la question de la «race» et semble être totalement autiste aux faiblesses des thèses qu’elle défend…..
Aussi darwincwg, je t’invite à te renseigner davantage en matière d’hérédité, de génétique des populations, de biologie de l’évolution et de paléoanthropologie via des sources autres que l’équipe de joyeux drilles racialistes auxquels tu te réfères. Car vois-tu si les thèses racialistes de Rushton, Jensen, Lynn et compagnie ne sont pas considérés comme pertinentes par la majorité des anthropologues et des biologistes ce n’est pas pour des raisons politiques mais bel et bien pour le manque total de bases et de rigueur scientifiques des thèses en question.
En espérant que tu tiendras cette fois-ci des arguments qui te sont présenté comme il se doit.
À bon entendeur!
Salut!
Références:
[1] Bertrand Jordan (2008), L’humanité au pluriel, Éditions du Seuil 2008
[2] Peter A. Underhill et al (2000), Y chromosome sequence variation and the history of human populations, Nature
[3] Heng Li and Richard Durbin (2011), Inference of human population history from individual whole-genome sequences, Nature
[4] Henrik Kaessmann Victor Wiebe and Svante Pääbo (1999), Extensive Nuclear DNA Sequence Diversity Among Chimpanzees, Science
[5] Anne C. Stone et al (2002), High levels of Y-chromosome nucleotide diversity in the genus Pan, Proceedings of the National Academy of Sciences
[6] Anne Fischer et al (2003), Evidence for a Complex Demographic History of Chimpanzees, Molecular Biology and Evolution
[7] Pascal gagneux et al (1999), Mitochondrial sequences show diverse evolutionary histories of African hominoids, Proceedings of the National Academy of Sciences
[8] Jean-Jacques Hublin et Bernard Seytre (2008), Quand d’autres hommes peuplaient la Terre, Éditions Flammarion 2008
[9] Michael Bolus et Nicholas Conard (2011), Les débuts de la culture en Europe, Pour La Science, numéro 407 -Septembre 2011
[10] Jared Diamond (1992), De l'inégalité parmi les sociétés, Éditions Gallimard 1997
[11] Susanne M. Jaeggi et al (2008), Improving fluid intelligence with training on working memory, Proceedings of the National Academy of Sciences
[12] T. Steinger, P. Haldimann, K . A. Leiss and H. Müller–Schärer (2002), Does natural selection promote population divergence? A comparative analysis of population structure using amplified fragment length polymorphism markers and quantitative traits, Molecular Ecology
[13] Willerman, L., Naylor, A. F., & Myrianthopoulos, N. C. (1974), Intellectual development of children from interracial matings: Performance in infancy and at 4 years, Behavior Genetics
[14] Un plaidoyer et une mise en garde contre le QI, Le Journal des Psychologues, numéro 230 – septembre 2005
[15] Ned Block, Race, gènes et QI, La Recherche
[16] Barbara Tizard et al (1972), Environmental Effects on Language Development: A Study of Young Children in Long-Stay Residential Nurseries, Institute of Education, University of London
Inscription à :
Articles (Atom)